
Neural Control of Movement
Cerebellum
The cerebellum (CB) connects the higher levels of the motor control hierarchy with intermediate and lower levels. The CB is important for the integration of sensory with motor signals – it receives input from all sensory systems and its outputs target all structures involved in the generation of movement.
Gross anatomy
The cerebellum consist of cerebellar cortex (CBX) and deep cerebellar nuclei (DCN). The cerebellar cortex is convoluted into many folia. Three major transverse divisions – lobes – are recognized. The anterior lobe is most rostral, the posterior lobe and flocculo-nodular lobe are located more caudally. These lobes are divided by the primary fissure and the posterolateral fissure, respectively.
Independent of the transverse, lobular arrangement, three major sagittal divisions are recognized. The central longitudinal strip of cortex is the vermis. On each side, the vermis is divided from the lateral cerebellar hemispheres by the narrow pars intermedia. In humans, the hemispheres are so highly developed that they obscure pars intermedia and much of the vermis.
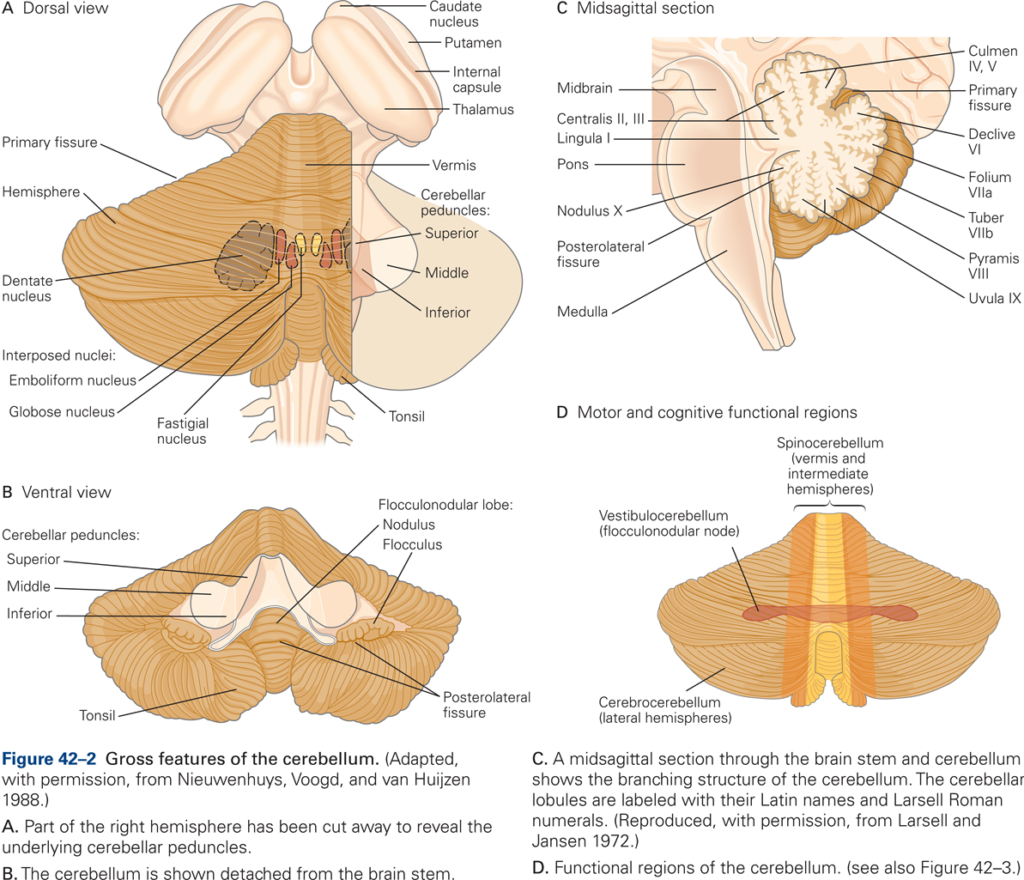
All input to and output from the cerebellum are via three peduncles on each side. All output from the cerebellum, via the deep nuclei, is through the superior cerebellar peduncle (brachium conjunctiva). The middle cerebellar peduncle (brachium pontis) is exclusively input from the pontine nuclei. The inferior cerebellar peduncle (restiform body) contains exclusively inputs from the spinal cord and brainstem including, in particular, the inferior olive.
Functional organization
The cerebellar cortex overlies a core of white matter. At the centre is a collection of nuclei – the cerebellar deep nuclei (DCN). These are the output areas of the cerebellum, receiving inputs from the cortex and projecting out to the thalamus, red nucleus and brainstem. From medial to lateral they are the fastigial, interpositus (in man, this is subdivided into the globose and emboliform nuclei) and dentate nuclei.
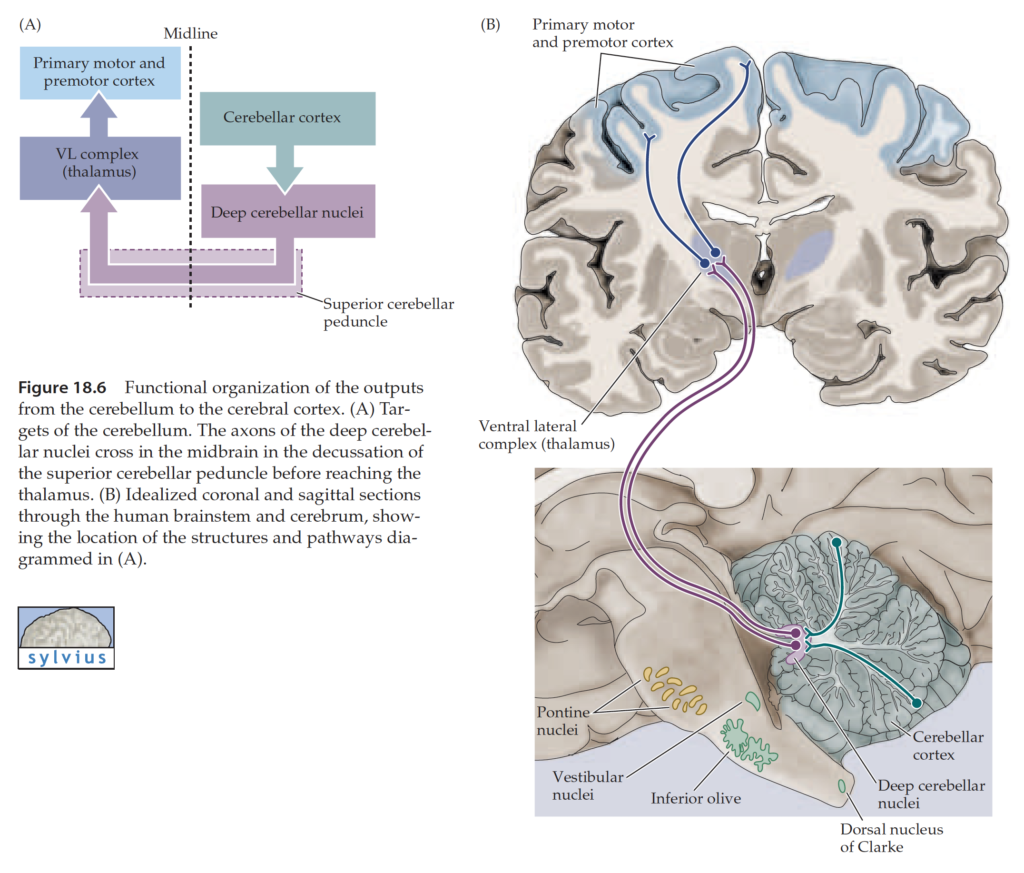
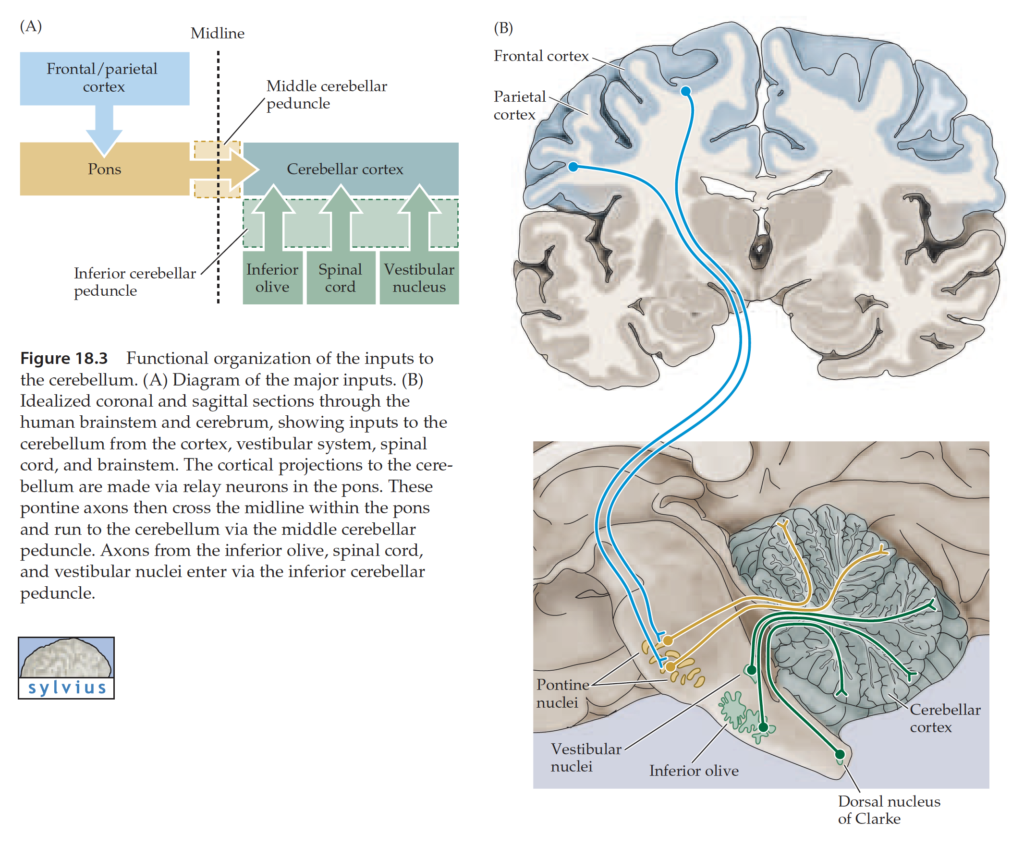
The cerebellum receives two types of afferent inputs: climbing fibers (CF), which originate from the inferior olivary nucleus in the medulla, and mossy fibers (MF), which originate from multiple sources (spinal cord, pontine nuclei (PN), reticular formation, vestibular nuclei). Input fibers synapse on neurons in both CBX and DCN. The bulk of CBX output neurons (Purkinje cells) project first to DCN rather than directly out of CB.
Older studies of cerebellar connections suggested three major divisions of cerebellum based on a rough segregation of inputs. The flocculo-nodular lobe (phylogenetically oldest – archicerebellum) receives many vestibular inputs – hence vestibulocerebellum. The rostral and caudal parts of vermis (paleocerebellum) receive many spinal inputs – hence spinocerebellum. The cerebellar hemispheres and lobules VI and VII of the vermis (phylogenetically most recent – neocerebellum) receive many pontine inputs – hence pontocerebellum (cerebrocerebellum).
Each of the major sagittally oriented cerebellar subdivisions are compartmentalized into a series of parasagittal strips (for review see: Voogd and Glickstein 1998). Later studies have confirmed that cerebellar cortical afferents and efferents travel within these strips, or zones and, therefore, function relatively independently from one another.
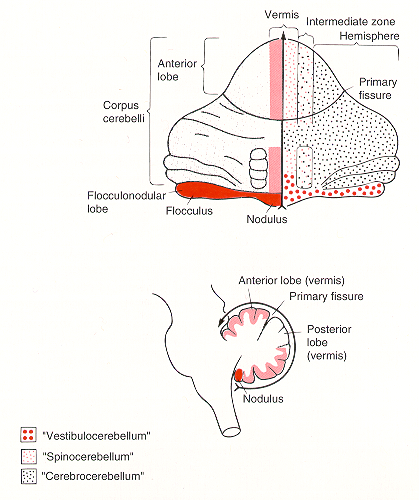
Vestibulocerebellum (flocculonodular lobe)
Inputs derive directly from primary vestibular afferents (semicircular canals and otolith receptors) and indirectly from secondary afferents that arise from the vestibular nuclei. Main output targets are the vestibular nuclei. The main functions of the vestibulocerebellum are to 1) control of axial and proximal limb muscles that are used to maintain balance, and 2) control eye movements and coordinate movements of the head and eyes (vestibulo-ocular reflex).
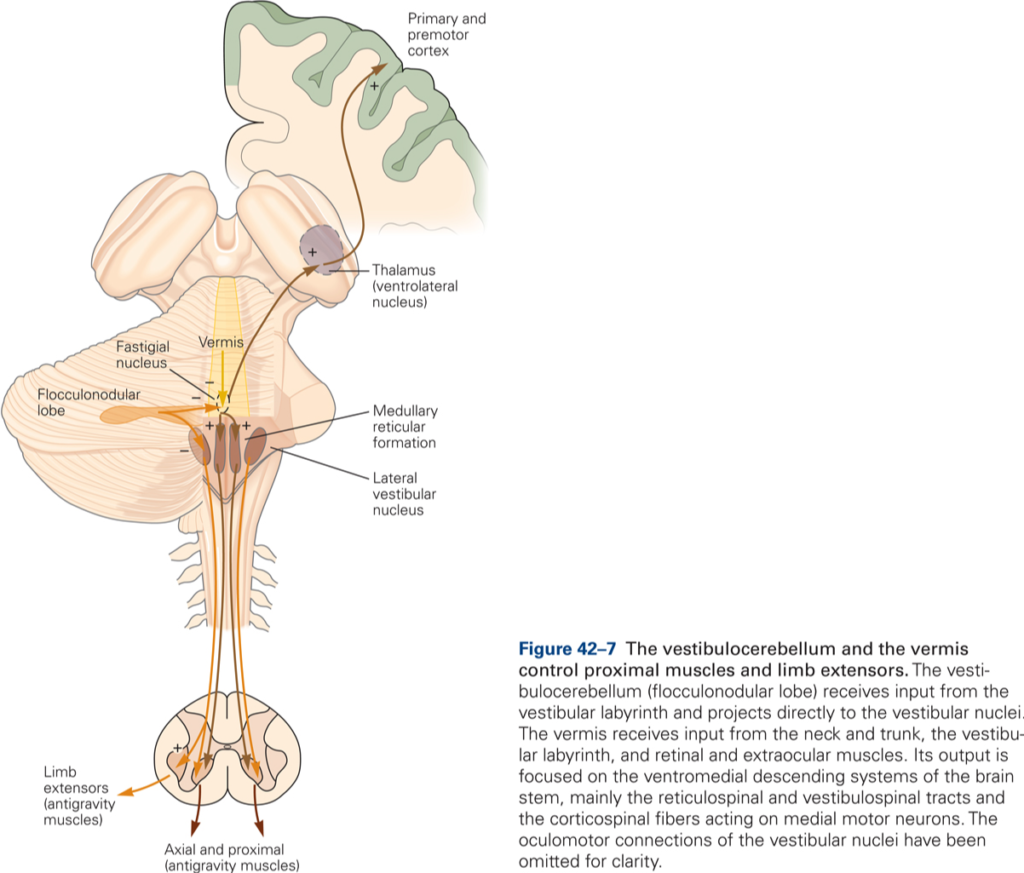
The main body of the CB is divided into three longitudinally arranged subdivisions: medial, intermediate, and lateral. Together the medial and intermediate subdivisions are referred to as the spinocerebellum, the lateral subdivision is called the cerebrocerebellum.
Spinocerebellum
Medial and intermediate zones receive somatosensory information from spinal cord via spinocerebellar tracts: the dorsal and ventral spinocerebellar tracts (DSCT and VSCT, respectively) are the pathways from the trunk and legs; the cuneocerebellar tract (CCT) and rostral spinocerebellar tract (RSCT) from the arms and neck. The DSCT arises from cells of Clark’s column in the lumbar spinal cord; the CCT originates from cells in the external cuneate nucleus in the medulla. Neurons in Clark’s column and in the external cuneate nucleus receive direct input from muscle spindles, GTO, joint and cutaneous receptors. DSCT and CCT transmit information about current limb position. VSCT and RSCT transmit information about the activity of spinal interneurons (such as the group Ia and Ib interneurons, which integrate descending and peripheral inputs). Intermediate zone receives additional inputs from motor cortex via the PN.
Medial zone projects via fastigial nucleus to cortical and brainstem components of the medial descending system (e.g. ventral corticospinal, reticulospinal, and vestibulospinal tracts), which influence axial and proximal muscles.
Intermediate zone projects via interpositus nucleus to the cortical and brainstem components of the lateral descending system (rubrospinal and lateral corticospinal tracts), which influence distal limb muscles.
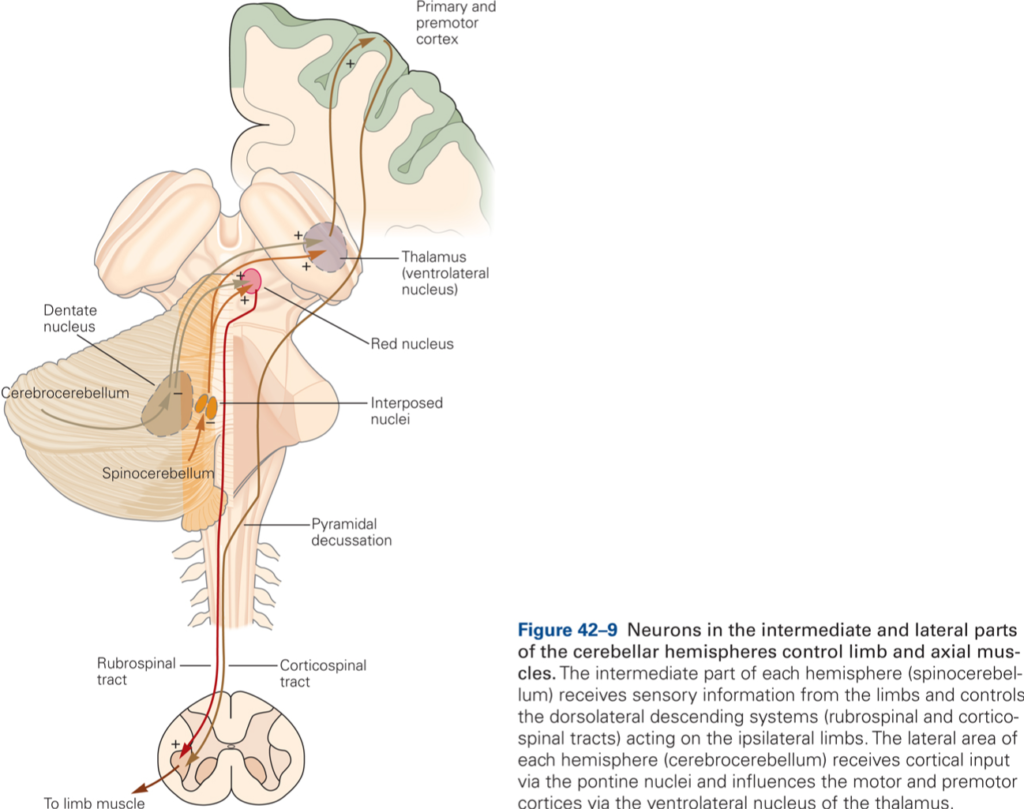
Spinocerebellum controls execution of movement and regulates muscle tone. Intermediate CB compares the actual limb position as provided by spinocerebellar input with the desired limb position provided by information about movements from higher brain centers, such as motor cortex. When actual and desired limb position differ (for example because variations in load are encountered during the movement), intermediate CB issues signals that are used to minimize the mismatch. These signals are transmitted via the rubrospinal and corticospinal tracts. Intermediate CB is involved with the detailed execution of movement.
Cerebrocerebellum
Input to cerebrocerebellum derives from sensory and motor cortices and premotor and parietal cortices (via the PN).
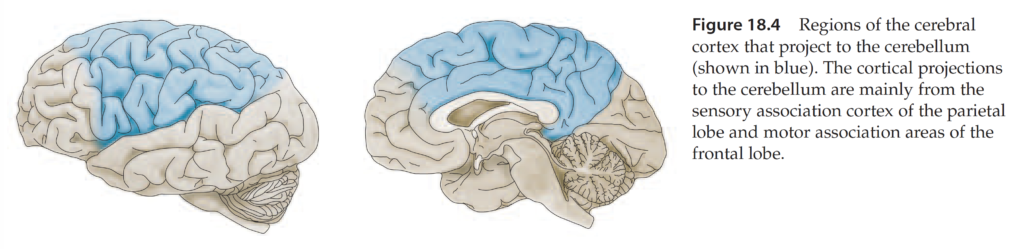
Output of cerebrocerebellum derives from the dentate nucleus and targets (via VL thalamus) motor and premotor areas of cerebral cortex. Major function of cerebrocerebellum is in the planning and sensory (especially visual) guidance of movement. The lateral cerebellar zone receives visual information appropriate for guidance of movement from posterior parietal cortex (PPC). PPC relates somatosensory information about limb position with visual information about location of objects in extrapersonal space.
How does vision guide movement, that is, how does visual information reach motor cortical areas? Visual signals reach motor cortical areas not via corticocortical connections, but via the following pathway: PPC –> PN –> lateral CB –> dentate nucleus –> VL thalamus –> motor and premotor cortices.
Lateral CB compares target location with limb position. Using continuous visual feedback to compare target location and limb position is not the preferred way to provide visual guidance because this process is slow and fast out of date with moving stimuli. Lateral CB is involved in predictive control, its signals predict where the limb is going to be based on how fast the target is moving.
Clinical signs of cerebellar damage
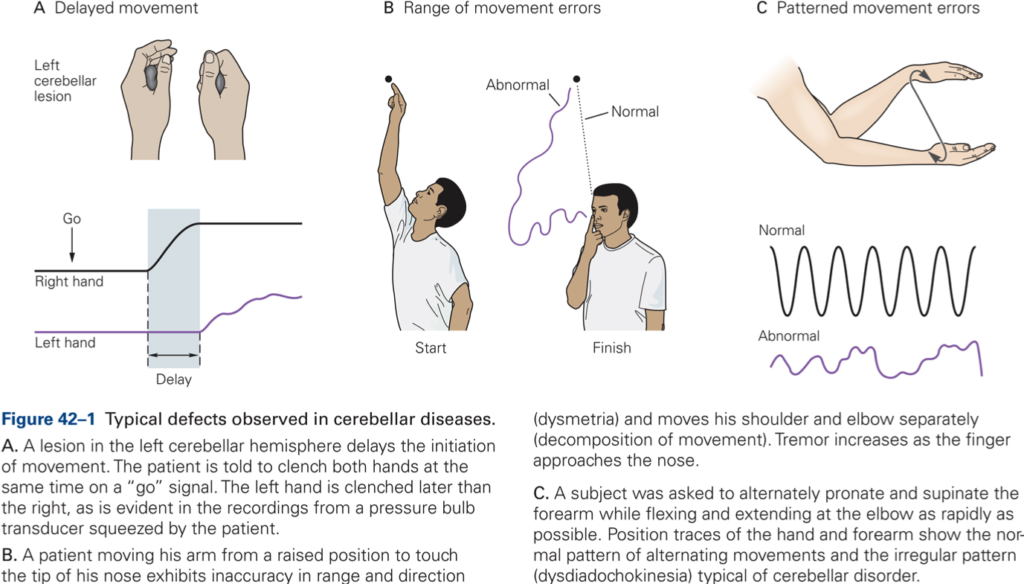
Ataxia
Syndrome characterized by a breakdown of movement coordination and inaccuracies when reaching for a visual target. Ataxia includes:
- Dysmetria (inaccurate movement amplitudes)
- Asynergia (poor coordination between joints, decomposition of movement)
- Intention tremor
- Dysdiadochokinesia (inability to perform rapidly alternating movements smoothly)
- Hypotonia and pendular knee jerks.
Localization of function (somatotopic organization)
Lesions in intermediate and lateral CB give rise to ataxia of individual limbs; lesions in medial CB results in poor equilibrium and falling, ataxia of gait (walking).
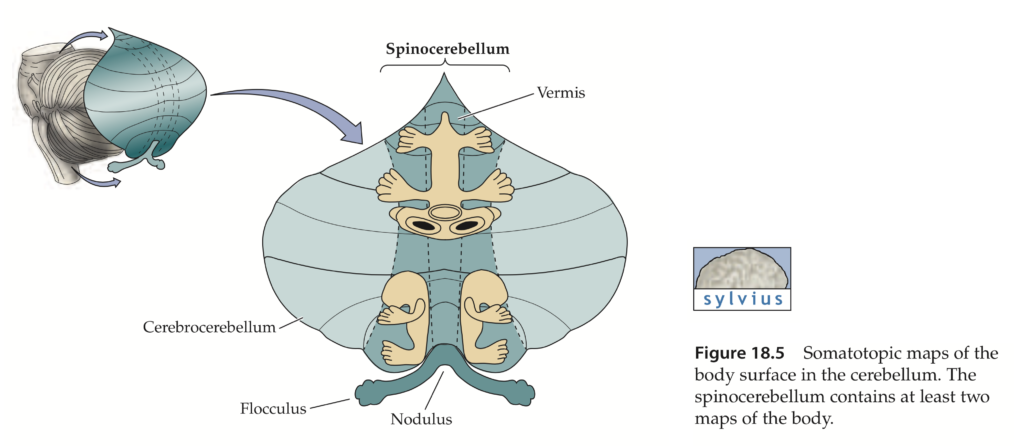
CB damage prevents modification and adaptation of behaviors, that is, it has a function in motor learning.
Cerebellar cortical circuitry
The cerebellar cortex is composed of 5 main neuron types. These are arranged with perfect geometrical precision throughout the cortex. One area of cortex is indistinguishable from another. Although not yet understood, it is widely held that the key to understanding the cerebellum is to understand the relationship between mossy and climbing fibre inputs.
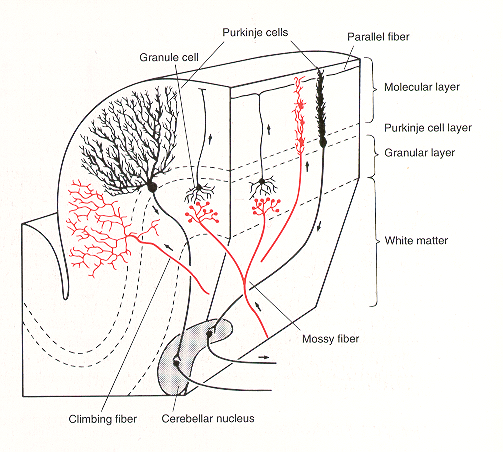
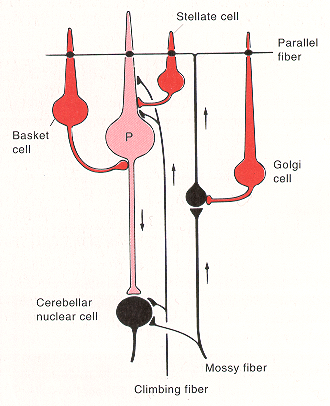
- Purkinje cells – large pyramidal cells. Output cells of cortex, inhibitory upon the deep nuclei. Their soma lie in the Purkinje cell (PC) layer and their flat fan-shaped dendrites project upwards into the molecular layer and are oriented in a plane perpendicular to the long axis of the cerebellar folia. Transmitter – GABA.
- Granule cells – most numerous cells in the whole brain. Receive mossy fibre inputs and give off an ascending axon that rises to the molecular layer, splits to a T-shape to give two parallel fibers (PF). PFs run along the long axis of lobules, perpendicular to PC dendritic trees. Each terminates on one or more PCs. Each PC receives excitatory inputs from up to 200,000 PFs. Transmitter – glutamate.
- Climbing fibers – from the contralateral IO. Each climbing fibre contacts only one PC but makes multiple synaptic contacts with it – the most powerful synapse in the nervous system. Transmitter glutamate? or aspartate?
- Golgi cells – morphologically slightly similar to PC but their inhibitory outputs are within the cortex – to granule cells. Transmitter – GABA.
- Basket and Stellate cells – receive PF inputs and are inhibitory on PCs – transmitter GABA?
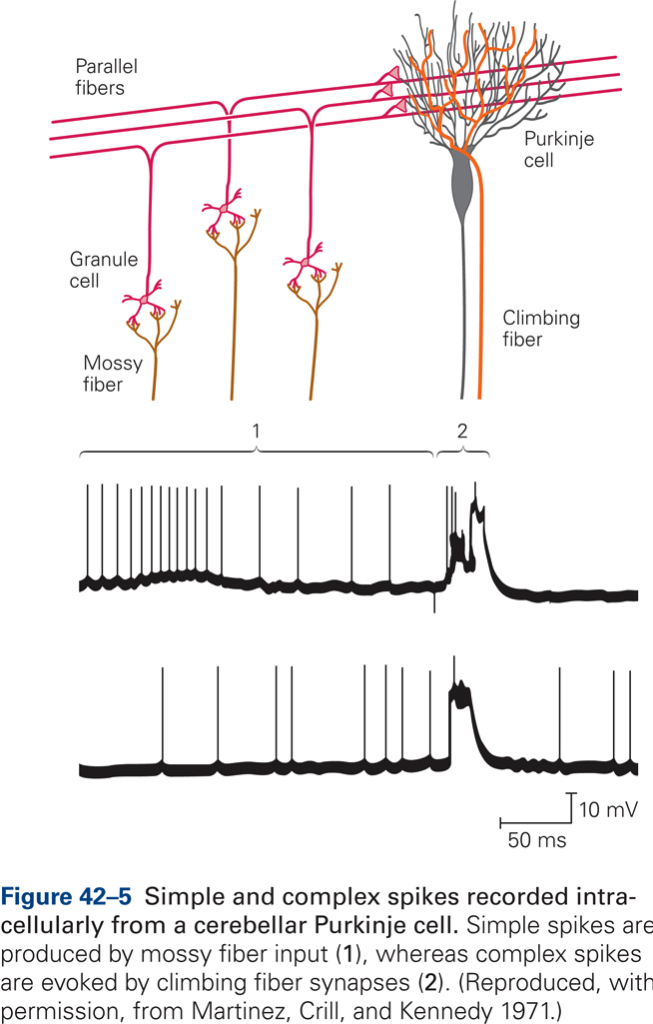
The synapse between CF and P cell is extremely powerful: a single action potential in the CF produces firing of the P cell (complex spike). CF fire at low rates (~1 spike/s). In contrast to CF, MF fire action potentials at high rates and are thought to transmit detailed parameters of movement. MFs generate simple spikes in PCs.
Motor learning
Motor learning is defined as a set of processes associated with practice or experience leading to relatively permanent changes in skilled behavior.
Cellular basis of motor learning
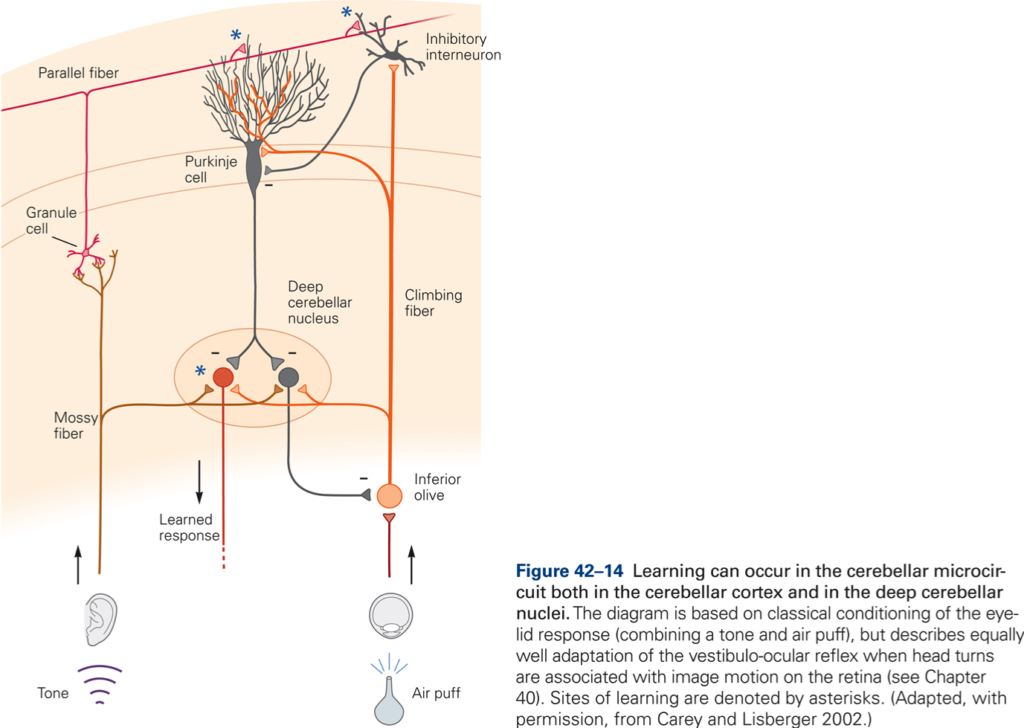
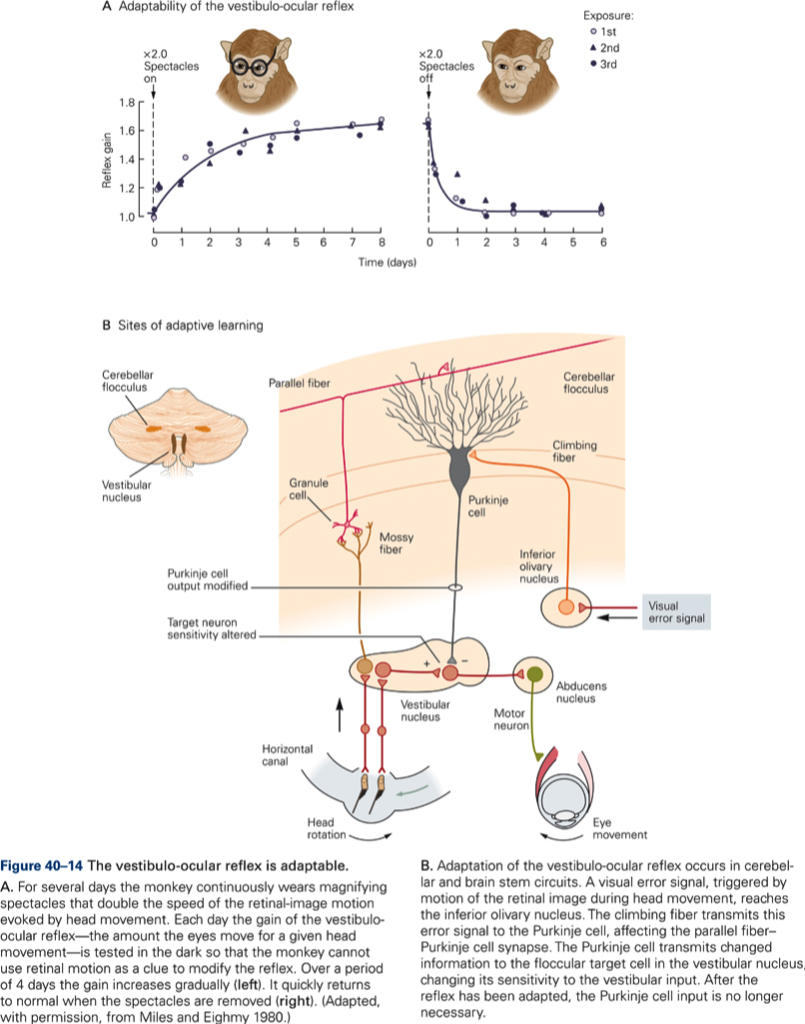
The central postulate in any theories of a role of CB in motor learning is that the activity in CF can modulate the effectiveness of the PF – P cell synapse (CF ‘teaches’ the cerebellar cortex (CBX) how to respond to particular patterns of PF inputs. Thus during the learning period we would expect an increase in CF activity, which should decrease once learning is complete. Over the same period, simple spike activity of P cells evoked by PF input should change (increase if CF increase synaptic efficacy) and should remain at a new value after CF activity had returned to normal –> a long-term change in synaptic effectiveness of PF is produced by a short-term change in CF discharge. Evidence for such a scheme has been obtained from recording experiments in monkey. Additional evidence for a role of CB in motor learning is derived from studies of adaptation of the vestibuloocular reflex (VOR) and from studies using classical conditioning of eye blink reflex. CB damage or lesions prevent VOR adaptation and prevent learning and retention of conditioned response.
https://mediaspace.wisc.edu/id/1_uth5tkvf?width=649&height=401&playerId=25717641
Kaltura Mediaspace
Although it is generally agreed upon that CB plays a role in motor learning, there is much debate about whether CB is the primary site of learning or whether it is necessary for learning to occur and the ‘memory’ is stored somewhere else. In other words, are the synaptic changes which occur during learning in the cerebellum?
References
- Voogd, J., & Glickstein, M. (1998). The anatomy of the cerebellum. Trends in Neurosciences., 21(9), 370-375. [PDF]
- Glickstein, M. (2000). How are visual areas of the brain connected to motor areas for the sensory guidance of movement? Trends in Neurosciences., 23(12), 613-617. [PDF]
- Glickstein, M. (2007). What does the cerebellum really do? Current Biology, 17(19), R824-827. [PDF]
