
M1 | Neuronal Mechanisms for Information Transmission
Neural Integration
Key Takeaways
Complex integrative functions arise from a limited number of basic neuronal interactions such as:
- Summation
- Facilitation/disfacilitation
- Inhibition/disinhibition
These fundamental processes operate during communication between groups of neurons as well as in the communication of one neuron with another. The ways in which neurons are interconnected is modifiable (plastic) during development, and later through learning. Neuronal plasticity underlies individuality: each one of us is unique.
Simple integration
Summation of excitatory postsynaptic potentials (EPSPs) and inhibitory postsynaptic potentials (IPSPs) at synapses. The final output of the neuron, an action potential, represents how that neuron encoded all incoming EPSPs and IPSPs.
Fundamental neuronal interactions
Facilitation
![]()
Facilitation involves excitatory synaptic interaction. So, when an AP invades the presynaptic terminal, Ca channels open, and the resulting increase in Ca2+ concentration determines how much neurotransmitter is released in the synapse which, in turn, determines the amplitude of EPSPs in the postsynaptic neuron. At low AP frequencies, the increase in [Ca2+] associated with any single AP has returned to baseline before the next AP invades the terminal. However, at higher AP frequencies, a burst of APs (rather than a single AP) invade the presynaptic terminal, which results in a cumulative increase in [Ca2+], and thus a cumulative increase in EPSP amplitude, with each subsequent AP. This process, that output increases with each subsequent AP, is referred to as facilitation: higher frequencies of action potentials release more neurotransmitter and thus produce larger amplitude graded potentials. Facilitation may last for seconds and therefore may contribute to longer term storage of information and to changes in responses during learning. In its simplest form, stored information here refers to the level of activity of the presynaptic neuron that immediately precedes the occurrence of each individual AP.
- Neuron becomes more responsive to its input (simple form of learning).
- Mechanism: accumulation of Ca2+ in presynaptic terminal —> subsequent action potentials release more transmitter —> increased Na+ conductance in postsynaptic membrane —> higher firing rate in postsynaptic neuron (red).
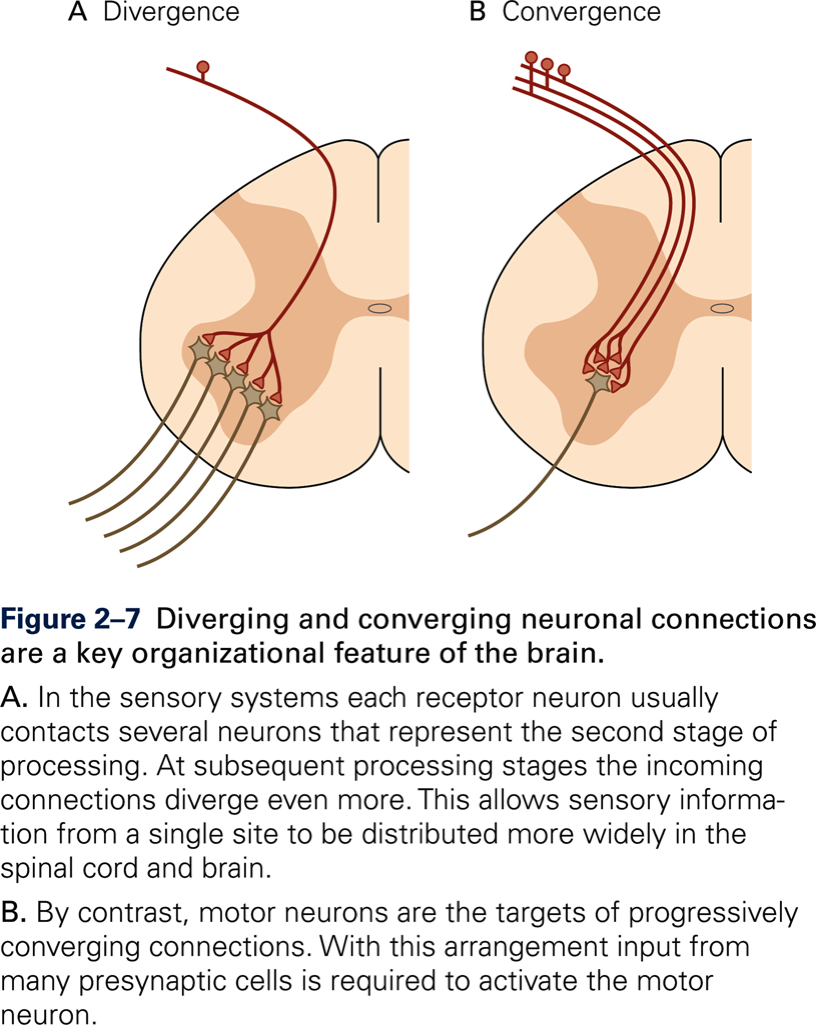
Disfacilitation
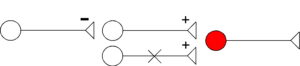
- Inhibition by neuron (on left) results in decreased neuronal activity of output neuron (in red), through inhibition of facilitatory (excitatory) interneuron (middle).
- Mechanism: decrease of Na+ conductance in membrane of postsynaptic neuron (red).
The neuron with the X through it (bottom) illustrates the overall effect on the post-synaptic, output neuron (in red). Here, during disfacilitation, the X refers to removing facilitation, thus reducing output of the circuit by decreasing activity of the post-synaptic neuron (red). During disfacilitation (top), facilitatory influences by neuron 2 (middle) on the output neuron (right, in red) are reduced through inhibition, exerted by neuron 1 (left) on neuron 2 (middle).
Inhibition
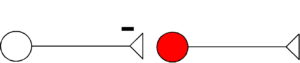
- IPSPs decrease probability that postsynaptic neuron fires an action potential.
- Mechanism: increase in membrane permeability for Cl– and K+ in post-synaptic neuron (red), resulting in increased hyperpolarization.
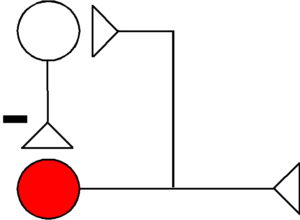
Disinhibition
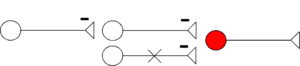
- Inhibition by neuron (on left) results in increased neuronal activity of output neuron (in red), through inhibition of inhibitory interneuron (middle).
- Mechanism: decrease of Cl– and K+ conductance in postsynaptic neuron (red), thus removing hyperpolarization.
The neuron with the X through it (bottom) illustrates the overall effect on the post-synaptic, output neuron (in red). Here, during disinhibition, the X refers to removing inhibition, thus increasing output of the circuit by increasing activity of post-synaptic neuron (red). During disinhibition (top), inhibitory influences by neuron 2 (middle) on the output neuron (in red) are reduced through inhibition, exerted by neuron 1 (left) on neuron 2 (middle).
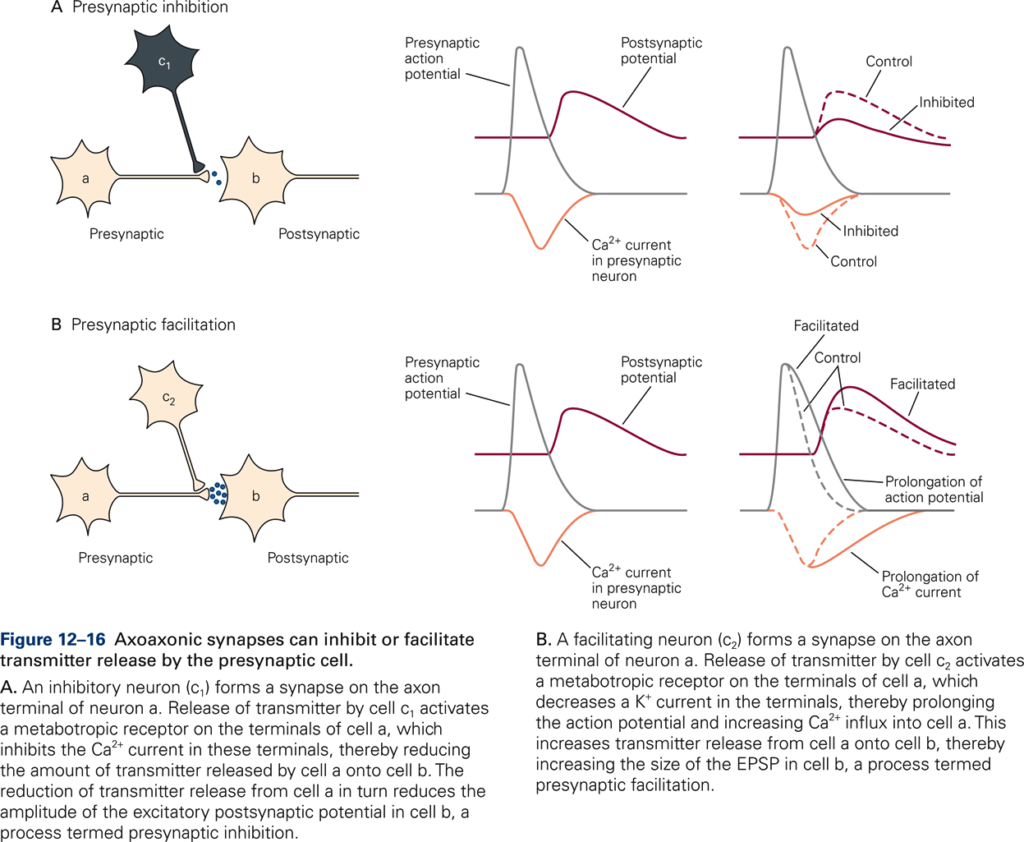
Presynaptic inhibition
- An inhibitory neuron c1 (via an axo-axonic synapse on a second cell a) reduces the amount of transmitter released by that cell on a third cell b.
- Mechanism: increased conductance to Cl– —> less depolarization produced by action potential —> fewer Ca2+ channels opened by action potential.
Example: Descending motor pathways transmit motor signals that produce movements. These motor signals inhibit (reduce) sensory input that is not needed in the production of the movement via presynaptic inhibition.
Presynaptic facilitation
- Increased influx of Ca2+ —> increase of transmitter release with each action potential.
- Mechanism: serotonin closes K+ channels (via 2nd messenger) —> longer duration action potential —> increased Ca2+ influx.
Central processing accomplished by circuits
The divergence and convergence of neuronal connections represent a key principle in the organization of the brain. In the sensory systems the receptor neurons at the input stage usually branch out and make multiple connections with neurons that represent the second stage of information processing. Subsequent connections diverge even more. By contrast, motor neurons are targets of progressively converging connections. With convergence, the target cell receives the sum of information from many presynaptic cells.
Divergence
- Serves to amplify or distribute effects: one neuron contacts many others, directly via axon collaterals, and indirectly via other neurons, which each contact many others.
- Predominantly present at input stages. For example, stretch of muscle activates hundreds of sensory neurons that in turn activate every motor neuron that innervates that muscle.
Convergence
- Serves to integrate information from many sources: one neuron receives connections from many presynaptic neurons.
- Predominantly present at output stages. For example: motor neurons receive multiple inputs from different types of sensory receptors and from descending motor pathways —> final common path
Recurrent inhibition
- An axon collateral of an active neuron projects to an inhibitory interneuron that then in turn projects back to the same neuron (feedback circuit). Serves to limit the excitatory response of the neuron by decreasing duration of bursts of action potentials.
- Example: axon collaterals of motor neurons activate Renshaw cells, which in turn inhibit motor neurons.
Recurrent facilitation
- Prolongs output. Pacemaker or reverberating circuits.
- Example: circuit between nucleus interpositus (NI), red nucleus, and lateral reticular nucleus, and NI.
Rhythmic alternating activity
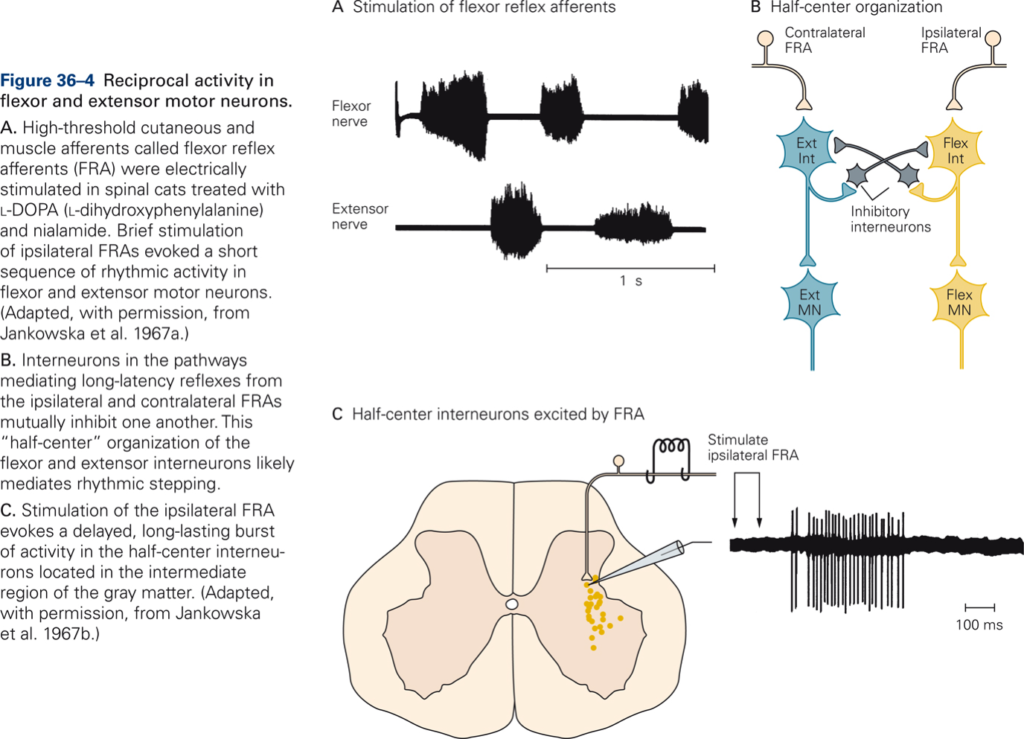
[Principles of neural science. (2013). (E. R. Kandel, J. H. Schwartz, T. M. Jesell, S. A. Siegelbaum, & A. J. Hudspeth Eds. Fifth ed.). New York: McGraw-Hill Medical]
- Half-center model. Flexor and extensor motor neurons with their associated interneurons each constitute one half-center. The two half-centers mutually inhibit each other, so that when one half-center is active, the other is inactive. It is assumed that switching between active half-centers occurs because the duration of inhibition by the active half-center is limited.
- Example: neural control of walking, chewing, breathing.
